THE PERMIAN WORLD
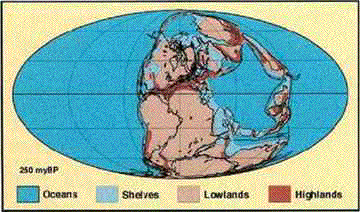 |
| The world of the Late
Permian. Present-day continents
are shown in outline: in the Southern Hemisphere (left to right),
South America lies against Africa, with India to the right. Below
are Antarctica, and to its right, Australia. Together these pieces
(South America, Africa, India, Antarctica, and Australia) form
Gondwana. In the Northern Hemisphere (left to right),
North America is adjacent to Western Europe, though much of Europe
is underwater. Siberia is further right. Together, they (North
America, Europe, Siberia) form Laurasia. Pangea
includes both Gondwana and Laurasia. The North
and South China blocks are the large masses in the ocean, well
to the right of Gondwana and Laurasia. The continents and the
China blocks enclose the Tethys Ocean; surrounding Pangea
is Panthalassa, the world ocean. The light blue color
refers to continental shelves, underwater parts of the continents.
(Map from Scholle, 1995, based on the work of Scotese
and others, 1979.) |
A map of the Earth at the time in Permian
time bears no resemblance to that of the present Earth. This is
because continents have been moving back and forth across the
surface of the planet, colliding to form mountain ranges, and
breaking apart to form oceans, ever since the molten rock of Earth's
earliest days first cooled sufficiently to allow large masses
of rock to congeal. (This probably occurred about four billion
years ago, some half billion years after Earth's formation, though
the timing of the origin of the continents is disputed.) For further
information on this process, see the Primer on Plate Tectonics:
APPENDIX
1: MOVING THE CONTINENTS; CREATING THE OCEANS
At times the continents that existed
have been relatively dispersed across the face of the globe, as
they are today. At other times, they have amalgamated into supercontinents,
containing the equivalent of several of today's continents. On
occasion, there have even been megacontinents, which comprise
in one continuous mass almost all the continental material on
the Earth's surface. Such was the case in the Permian, when a
major feature of global geography included a global megacontinent
we call Pangea, that is, "all-the-earth."
This megacontinent was comprised of two
immense landmasses ("supercontinents"), one composed
of what is now Antarctica, Australia, the Indian subcontinent,
Madagascar, Africa and South America and called Gondwanaland,
after part of India, Gondwana, the "land of the Gonds"
[so Gondwanaland is actually redundant, meaning land of the Gonds
land]. (The Gond kingdoms of Madhya Pradesh -- the "Middle
Province" of central India -- flourished from about the 12th
century on, and Gondwana was a powerful state in the 16th century.)
The other immense landmass consisted of what is now North America,
Greenland, Europe and a large part of Asia, and is referred to
as Laurasia. This name derives from a combination of the Laurentia
(northeastern Canada), geologically the most ancient portion of
North America (and often referred to as the Laurentian or Canadian
Shield) with Asia.
These two landmasses partially enclosed
a large tropical ocean, centered on the equator, called the Tethys.
Bounding the Tethys to the east were the North China and South
China blocks, great islands somewhat like today's Greenland or
New Guinea. Within the Tethys, north of Gondwana, was a string
of large islands (comprising today's Turkey, Iran, Afghanistan,
and Malaysia) that were part of the largely submerged Cimmerian
continent. (The Lhasa Block, a large island that eventually became
much of Tibet, may have been part of Cimmeria, or may have followed
it: Wignall and Twitchett, 2002.) The Cimmerian continent had
detached from Gondwana earlier in the Permian (or perhaps the
Late Carboniferous), and had begun moving across the Tethys. The
splitting of Cimmeria from Gondwana was caused by the development
of the "Tethian rift," a mid-ocean ridge, which was
eventually to transport Cimmeria all the way across the Tethys
and attach it to Laurasia. (The Tethian rift, because it produced
ocean floor, actually created a new ocean, labeled the Neo-Tethys,
and helped destroy an older one, called the Paleo-Tethys, which
subducted beneath Laurasia.)
Surrounding the entirety of Pangea as
well as the North and South China blocks was an immense ocean,
a hemisphere-sized ocean (therefore larger than today's Pacific):
the "all-ocean," Panthalassa. This was the largest ocean
of the Phanerozoic. Some paleoceanographers (those who study ancient
oceans) believe that Panthalassa was a stagnant, highly stratified
ocean, perhaps largely anoxic (lacking in the dissolved oxygen
needed by all animals and other organisms): "dead,"
except for its upper reaches (Wignall and Twitchett, 2002). The
evidence for this anoxic, stratified ocean, however, comes from
a single locality (Isozaki, 1994; 1997a), and that source has
indicated that the anoxia may have been the consequence of events
at the end of the Permian (Isozaki, 1997b), rather than having
been an ongoing condition of this ocean. Certainly careful studies
of the organisms in the ocean waters off the northwest coast of
Pangea reveal no such anoxia before the end of the Permian (Beauchamp
and Baud, 2002).
The Tethys, with just a few relatively
narrow passages (up to about 1000 kilometers/600 miles wide) connecting
it to the larger ocean (Panthalassa), may have had more restricted
circulation, although today's Mediterranean, with only a single
outlet through the Strait of Gilbraltar, exchanges large quantities
of water with the adjoining Atlantic. The evidence for anoxia
in the Tethys, at least during the last million years or so prior
to the end of the Permian (Wignall and Hallam, 1992; Twitchett
and Wignall, 1996; Wignall, 1995; Kershaw, 1999) is somewhat better
than for Panthalassa. Being a warm and largely enclosed ocean
would have made the equatorial Tethys more susceptible to oxygen
depletion (warm water holds less dissolved gas) than Panthalassa,
which stretched from pole to pole, and undoubtedly was recharged
with oxygen in its colder regions. Before the last days of the
Permian, however, it seems likely that the Tethys was quite a
normal (unstratified, oxygenated) ocean, and that Panthalassa
also likely was so right to the very end.
Large landmasses produce extreme weather
in their interiors. Whereas coasts tend to have climate conditions
which are milder due to the moderating effects of nearby oceans,
continental interiors have temperatures which are both hotter
in summer and colder in winter than those on the seacoast. The
midwestern regions of Canada and US, for example, can get quite
hot in summer, producing violent thunderstorms and tornadoes.
The same areas can be brutally cold in winter, with temperatures
descending regularly into the minus thirties (°C, = minus
twenties °F) in Missoula, Montana and into the minus forties
(°C or °F) in Edmonton, Alberta. By contrast, Ketchikan,
Alaska, though further north but on the Pacific coast, never has
such extreme temperatures. The coldest spot on Earth during the
Northern Hemisphere's winter is frequently around Yakutsk in the
interior of Siberia, rather than places further north.
Large landmasses not only have climate
extremes in their interiors, however, they also generate weather
extremes on their periphery. The monsoons that bring seasonal
rains to southern Asia, for example, are largely the creation
of low pressure systems that form over the Asian continent. The
heat of summer in the interior of Asia warms the air, causing
it to rise and surface air pressure to drop. This low pressure
region pulls in moisture-laden air from surrounding oceans, drawing
major storms to the Asian coasts. Because of the great size of
Pangea, it seems likely that its monsoons were proportionately
greater than today's.
The surface layer -- about a hundred
meters (yards) deep -- of Panthalassa would have been well stirred
by such tempests, particularly near the shore. Wind generated
waves would also have helped mix the surface layer, bringing nutrients
to the surface and oxygen into the water. The size of waves on
a given body of water is the uninterrupted expanse over which
the wind can blow, a dimension known as fetch.
The reason that waves on a small lake
are smaller than waves on a larger lake, other things being equal,
is that the smaller lake provides less fetch for waves to build.
On Panthalassa, fetch would have extended for half the globe,
allowing wind waves to build to considerable heights. In addition,
great oceanic swells (ultimately the consequence of the rotation
of the Earth), known as Rossby waves, may pump nutrients from
deeper layers to the surface of the ocean and thereby feed photosynthetic
algae. The churning of the upper regions of the ocean by this
mechanism has been labeled "the Rossby Rototiller" (Siegel,
2001).
Toward the end of the Permian, two major
volcanic events took place. One was an episode of significant
pyroclastic volcanism in southern China. Pyroclastic volcanism
is a particularly violent kind of volcanism, exhibited today around
the "Ring of Fire," the lands which border the Pacific
Ocean. In the Andes of South America, the Cascades of western
North America, the Aleutian Islands of Alaska, the Kamchatka Peninsula
of the Russian Far East, in Japan, the Philippines, Indonesia,
and many volcanic islands of the South Seas, we regularly see
highly explosive volcanoes called stratovolcanoes producing devastating
eruptions of ash and lava.
In recent decades, we have seen such
eruptions at Mount St. Helens in the state of Washington, Mt.
Pinatubo in the Philippines, and Mt. Unzen in Japan. Stratovolcanoes,
when they are not in their eruptive state, are often beautiful,
symmetrical mountains, draped with evergreen forests on their
slopes and capped with brilliantly white fields of snow and ice.
When they erupt, however, they produce huge dark clouds of ash
and gas which rain destruction on nearby communities. Some of
these immensely hot ash and gas flows, called pyroclastic flows,
race along at speeds of greater than a hundred miles per hour,
hugging the ground, searing and suffocating all living things
they encounter. Stratovolcanoes also erupt blobs of molten lava
which fall as rocky bombs on areas close by. The heat produced
during the eruptions of stratovolcanoes can melt their snow and
ice fields, sending floods of water, mud, and rock (called lahars)
surging down nearby river valleys. These lahars may carry volcanic
debris a hundred or more kilometers (about 60 miles) from its
source.
Particularly violent stratovolcanic eruptions
can actually blast away the summits of their peaks, as happened
with Mount St. Helens. Even more violent was the eruption about
10,000 years ago of Oregon's Mt. Mazama, which blasted away most
of the mountain. Mount Mazama exists no longer; its rim forms
the retaining wall of the body of water we now call Crater Lake.
Yet more was the 1883 eruption of the Indonesian stratovolcano
Krakatoa. Tens of thousands of Indonesians perished in the tsunamis
it produced; the sound of its explosion was widely heard around
the world. Thousands of others in the Roman cities of Pompeii,
and Herculaneum died in the pyroclastic clouds unleashed in 79
CE during the eruption of the stratovolcano Vesuvius. The 1902
eruption of Mount Pelée on the Caribbean island of Martinique
killed almost 30,000 people, including the entire population of
the city of St. Pierre: when the pyroclastic clouds passed, a
single human being was left alive in the city prison.
Despite such violence, however, stratovolcanic
eruptions do not cause mass extinctions. They can cause climate
effects, but even those are limited and short-term.
The pyroclastic eruptions of South China
stratovolcanoes about 250 million years ago, therefore, are extremely
unlikely to either have caused the end-Permian extinction, or
even contributed in any significant way to that event. This is
because, despite the fact that stratovolcanoes usually exist in
long chains or arcs rather than as isolated volcanoes, they don't
erupt all at once. Usually one volcano in a chain will erupt,
then, some decades later, a different one. This is how the chains
of volcanoes in Indonesia, or the Andes, or the Cascades behave,
so our evidence is good. In addition, though each eruption does
cause significant harm and disruption to nearby living things
-- demolishing forests and their inhabitants, for example -- the
effects are generally quite local and the affected organisms are
likely to inhabit ecologically similar areas elsewhere in the
volcanic chain. The likelihood of more than a few species -- if
that -- going extinct in a major volcanic eruption, therefore,
is very minimal.
There is, however, a second kind of volcanism
that occurred about 250 million years ago which could have had
much greater consequences. Near the end of the Permian, in northern
Siberia, was the largest volcanic episode of the Phanerozoic.
Instead of being produced by ordinary volcanoes, the eruptions
likely occurred through great cracks or fissures, spilling huge
quantities of syrupy, basaltic lava out over vast areas of the
Siberian countryside.
Fissure eruptions are an uncommon type
of volcanic eruption, generally unfamiliar even to those knowledgeable
about other kinds of eruptions. The most recent large fissure
eruption took place in southeast Iceland in 1783-84. There the
Laki (Kagagigar) eruption ripped open a crack in the surface some
25 kilometers (15 miles) long, and sent hot, fluid basalt a distance
of some 45 kilometers (27 miles). The lava eventually covered
about 500 square kilometers (about 200 square miles; Duff, 1993,
p. 210-11).
 |
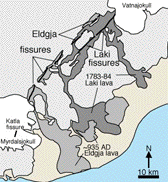
Laki fissure eruption. The map shows the areal extent of the eruption.
The photo looks southwest along the fissures, with a cinder cone
in front. (Duff, 1993, p. 210-11) |
The eruption produced fluorine-laced
ash, which contributed to crop failure and to the deaths of about
80% of Iceland's sheep population, largely from fluorine poisoning.
The ensuing famine and fluorine poisoning led to the starvation
of some 10,000 people, about 20% of Iceland's total population
(Fisher, 1997, p. 170; Thordarson and Self, 2003). An acidic atmospheric
haze created by the eruption reached the cities of northwestern
Europe, where it was observed as a "dry fog" by the
American ambassador to France, Benjamin Franklin (Brands, 2000,
p. 626). (A similar acid fog is apparently a feature of the Hawaiian
volcano Kilauea, where its acidity has created the adjoining Kau
Desert: Schiffman, 2006).
The Siberian Traps fissure eruptions
of 250 million years ago were accompanied by considerable pyroclastic
activity. About 20% of the material erupted by Siberian Traps
volcanism was in the form of pyroclastic debris, which produces
distinctive rocks called tuffs. Some of this pyroclastic volcanism
may have been of a particularly devastating variety called caldera
explosions, where an area up to perhaps thirty miles in diameter
suddenly settles into an enormous subterranean magma lake, lofting
ash high into the atmosphere and carrying it far from its source.
Ash from the Long Valley Caldera (California) explosion, which
happened about 730,000 years ago, distributed ash over about two-thirds
of what are now the contiguous forty-eight states. Caldera eruptions
associated with today's Yellowstone, going back some 17 million
years, regularly dropped ash over large parts of the country.
But fissure eruptions like those in Siberia
at the end of the Permian extrude much of their output as fast
flowing lava, much as do today's Hawaiian volcanoes. The thin,
syrupy basalt flows in sheets across relatively flat terrain,
eventually cooling into what are called "large igneous provinces"
or LIPs. Much of the Columbia River valley in Idaho, for example,
is occupied by such a LIP, providing farmers with fertile soil
for potato crops and tourists with the Craters of the Moon National
Monument.
LIPs are characterized by repeated, intermittent
basalt flows, which settle and cool on each other like layers
on a cake. These flows can look like giant steps, giving them
the Dutch name "Traps," meaning steps (Alvarez, 1997,
p. 168, fn. 24). In the northwest of India lie the famous Deccan
Traps, which erupted about 65 million years ago -- roughly the
same time as the end-Cretaceous extinction that killed the dinosaurs
-- and have helped confuse the issue as to that extinction's cause
(though it now seems to most scientists that the Deccan Traps
eruption played no role in that extinction). Similar LIP-produced
step-like formations may be found in Washington state's Grand
Coulee area.
 |
| A large igneous province
(LIP) in Greenland. The layers
in this flood basalt formation (presumably part of the widespread
North Atlantic Igneous Province, or NAIP) are horizontal, which
is the consequence of numerous thin, syrupy flows of basalt,
each having cooled over one previously deposited. (Duff, 1993) |
There is much about LIPs which we don't
know, because geology of necessity relies greatly on the principle
that "the present is key to the past." In other words,
geologists come to understand much of what occurred in the past
by seeing similar processes occurring today. But if a geologic
event is sufficiently rare (or even unique), it may be difficult
to understand all the consequences of that event. Thus, while
we can examine the consequences of the eruptions of ancient stratovolcanoes
because one or another present-day stratovolcano is always erupting
(actually, about twenty are erupting worldwide at any given time;
Simkin and Siebert, 2000, cited by Wright and Flynn, 2004), we
have no contemporary LIP eruptions to compare with the ones we
know took place in geologic time. Today's closest similar eruptions
may take place in Iceland, but as Iceland's volcanism is part
of the general mid-oceanic ridge system's extrusive activities,
it may not make for an appropriate comparison. Nonetheless, it
is clear that LIPs are produced by the eruptions of huge blobs
of mantle magma that have accumulated just below the crust of
the Earth.
|
Rising from the Deep,
1:
The Pennsylvanian-Permian Reversed Superchron, 312-262 Ma
It is possible, however, that the huge amount of material erupted
by the Siberian Traps was related to a strange occurrence in
the tens of millions of years that preceded the end of the Paleozoic.
The Earth's magnetic field, which compasses rely on to provide
an approximate direction for north and south, is actually quite
unstable over long periods of geologic time. Not only do the
positions of the magnetic poles move slowly about (a characteristic
known as polar wander), but they also occasionally and spontaneously
reverse direction: north becomes south and south, north. Nothing
happens to the planet during these times; the Earth itself does
not abruptly flip over, for example, but the polarity of its
poles suddenly shifts. The geophysicists who study such matters
trace these polarity reversals to random variations in the molten
outer core of the planet. (Those geophysicists who study the
history of Earth's magnetism are properly called paleomagnetologists,
but some of them enjoy labeling themselves paleomagicians.)
When lava erupts from
volcanoes or along the mid-oceanic ridge, the iron particles
in the lava, though initially pointing at random, orient themselves
toward the poles like tiny compasses. Upon cooling, they therefore
record in the rock the orientation of Earth's magnetic field
at the time. When the tiny iron compasses in the rock point in
the direction of today's north pole, the poles are said to exhibit
normal polarity; when they point south instead, the poles are
said to be reversed. Using sophisticated equipment, the periods
of normal and reversed polarity are clearly seen in the rock
record. Despite the use of the term normal to describe the current
polarity of the Earth's magnetic field, however, there is nothing
"abnormal" about reversed polarity: it is just as common
in the rock record as "normal" polarity.
Polarity reversals occur
every several thousand to many millions of years. When the periods
of magnetic stability are relatively short in length they are
referred to as chrons, but the few that lasted for extreme lengths
of time are called superchrons. Towards the end of the Paleozoic,
there was period when the magnetic polarity of the planet was
reversed for some fifty million years (from 312 to 262 million
years ago), the longest known period of magnetic stability of
the Phanerozoic. This interval is known as the Pennsylvanian-
Permian reversed superchron, for the two geologic periods during
which it occurred. (The Pennsylvanian Period is the younger portion
of the Carboniferous, which is divided into the Mississippian
-- about 360-320 Ma -- and the Pennsylvanian -- about 320-290
Ma.)
Something was clearly
somewhat different down in and near the planet's outer core.
We don't know what that something was, but it has been suggested
that it had to do with a disturbance at the boundary between
the outer core and the overlying planetary layer, the mantle.
It has been postulated that, on occasion, large blobs of mantle
rock, heated by the fierce heat of the core at the core-mantle
boundary, may rise through the mantle and cause eruptions on
the Earth's surface. (Some have even suggested that the material
at the bottom of the mantle, where the suggested blobs may originate,
may be the remnants of old tectonic plates that, once subducted,
ultimately drifted down that far.)
It is therefore possible
that a great blob of material accumulated at the core-mantle
boundary for the fifty million years of the Pennsylvanian-Permian
reversed superchron, stabilizing the planet's magnetic field.
When it became sufficiently buoyant, it lifted off and eventually
made its way to the planet's surface, where it erupted as the
Siberian Traps. This proposal thus explains both the existence
of the Phanerozoic's longest period of magnetic stability, why
it came to an end, and the origin of the Siberian Traps volcanism.
|
|
How We Know Where
the Continents Were
How do we know what
the configuration of the continents and oceans was like 250 million
years ago? One major way is by the use of paleomagnetism. As
lava is erupted, and iron-bearing particles orient themselves
according to the direction of the magnetic poles, those particles
record the position of the continent relative to Earth's magnetic
field. The iron partilces in each successive lava flow, upon
cooling, do the same. As a continent moves, therefore, the lava
flows from its volcanic eruptions record the position of the
continent at the time of eruption relative to the magnetic poles.
Using these "frozen compasses," it is possible to trace
the movement of a continent, or piece of continent, as it has
moved across the surface of the planet. The work is complicated
and difficult (magnetic reversals being one complicating factor),
but by carefully examining these natural compasses to determine
the original orientation of the ancient lava flows, scientists
can determine the ancient positions of continents -- or pieces
of continents.
Other evidence is also
used. Well before the discovery and use of paleomagnetism, some
geologists were struck by the similarity of fossils on continents
that are now quite distant from each other. Similar Permian-age
land plant fossils, for example, can be found in India, Madagascar,
South Africa, South America, Antarctica and Australia, all places
now separated by many thousands of kilometers (several thousand
miles) of ocean. To explain these similarities, geologists were
forced to posit some sort of physical connection (that would
allow the movement of terrestrial organisms) between these areas.
For years geologists
relied on the idea of "land bridges," like the famous
Bering Strait Land Bridge, across which the original human inhabitants
of the Americas crossed into the western hemisphere from Asia.
But unlike the Bering Strait Land Bridge, which really did exist,
and which was the result of the lowering of sea level during
the most recent ice age, there was no evidence for most other
proposed ancient land bridges. As we now know, large chunks of
land do not spontaneously pop up out of the ocean when needed
to allow the movement of terrestrial organisms from one place
to another, and then conveniently disappear beneath the waves.
Rather -- and probably even more amazing than the appearance
and disappearance of purported land bridges -- the continents
and pieces of continents themselves do move.
In the past, continents
which are now widely separated were actually joined together,
permitting the relatively easy movement of terrestrial organisms
from one place to another. The similar Permian plant fossils
of India, Madagascar, South Africa, South America, Antarctica
and Australia, for example, can now be explained by the fact
that for more than a hundred million years, these landmasses
were all part of the supercontinent Gondwana. Thus one line of
evidence of previous connections between continents is the similarity
of their fossils; another is the similarity of their rocks.
|
The Permian was the final geologic period
of the Paleozoic Era (543 to 250 million years ago). Named for
the medieval kingdom of Permia, which occupied a small region
between the Ural Mountains and the Volga River (Duff, 1993, p.
87) around what is now the city of Perm, Russia, where strata
from this period are prominent, the Permian lasted from about
290 until about 250 million years ago. In some ways, the creatures
of the Permian were beginning to resemble those of today. In other
ways, they were very different.
On the land, there were forests. These
forests were not like the forests of the preceding geologic period,
the Carboniferous, during which fern and lycopod trees occupied
swampy tropical lowlands and had begun to move into drier uplands.
(Though there are no longer any lycopod trees, lycopods are still
around, rising to the height of just a several centimeters (a
few inches) above wet forest floors. Referred to as club mosses,
they are not mosses at all, and an alternative common name, ground
pines, gives a better sense of what these plants look like.) These
trees became the source of much of the world's coal, conferring
the name Carboniferous upon the period.
The forests of the Permian were different.
Adapted to a drier world, Permian forests still provided the raw
materials for present-day coalfields, but their constituent trees
had changed. There are not any cycad trees around today, though
their unusual descendants, the cycads, are with us yet. In appearance
they are similar to ferns, though their fronds are much larger,
and generally tougher and drier. During the Permian, cycad trees
were a major component of forests, as were the conifers -- cone-bearing
trees that were the ancestors of today's conifers, pines, firs
and spruces.
Permian conifers apparently evolved in
the northern regions of Pangea, in the supercontinent called Laurasia,
and they had not yet spread to the southern supercontinent, Gondwana.
In Gondwana, the broad-leafed Glossopteris and its relatives were
the dominant trees. The Permian forests contained none of the
deciduous flowering trees of today's forests -- those trees hadn't
evolved yet -- but the northern coniferous forests would have
seemed vaguely familiar.
In the forests and on the plains of the
Permian world were vast numbers of a wide variety of reptiles,
which had largely replaced an earlier (Devonian-Carboniferous)
amphibian fauna. The giant insects of the Carboniferous -- millipedes
with bodies up to half a meter (a foot and a half) in length,
and dragonflies with three-quarter meter (two and a half feet)
wingspreads -- were gone. But there were numerous lineages of
reptiles.
Reptiles have a general competitive advantage
over amphibians. This advantage is not sufficient to drive amphibians
from every possible habitat: after all, there are still plenty
of amphibians in the world, hundreds of millions of years since
the Permian. Newts, salamanders, frogs and toads are quite plentiful,
but, of course, they all need water or highly moist conditions
in which to lay their eggs. But reptiles came up with an evolutionary
innovation which freed them from that requirement: the amniotic
egg. This egg, in essence, carried wet conditions with it. Having
a relatively hard shell, it was resistant to drying, and allowed
reptiles to move to areas like drier, upland forests, that were
generally inhospitable for amphibians.
During the Permian, some reptiles also
began to evolve a more efficient system of locomotion. Early reptiles,
like many amphibians, possessed sprawling limbs. But some reptiles
evolved the ability to place their limbs under their bodies, first
the hind limbs, then later, the front. The fully upright stance
was an evolutionary development that took many tens of millions
of years. But as the Permian progressed, having upright limbs
beneath the body became common, particularly among the more successful
lineages of reptiles. This position of the limbs permitted the
reptiles more rapid and efficient locomotion, increasing their
effectiveness as predators.
Many reptiles were not predators, however.
Unlike the amphibians, some reptiles developed the ability to
process large amounts of vegetation. More efficient jaw muscles
and more specialized teeth -- or, in some cases, horny beaks like
turtles -- allowed them to crop and chew plant matter. The girth
of some of these herbivorous reptiles testifies to the large guts
they employed for digesting large quantities of vegetation, and
their robust limbs indicate the physical strength required to
transport these heavy creatures around.
Another evolutionary innovation freed
many lineages of reptiles from obligatory cold-bloodedness. Most
reptiles, like many other organisms are cold-blooded, that is,
their body temperatures are just about the same as the world around
them. This limits their ability to be up and about during the
cooler hours of the day, and restricts them to warmer climates.
From the early Permian on, however, various lineages of reptiles
evolved solutions to this limitation. Most astonishing, perhaps,
were the sails that were developed by particular herbivorous and
carnivorous reptiles.
Labeled sails because that is what they
look like, these features were actually constructs of vertebral
bone, blood vessels, and skin that rose from the backs of reptiles
like Edaphosaurus (an Early Permian herbivore), and Dimetrodon
(a famous Early Permian carnivore). Most paleontologists believe
that the primary function of these sails (there may have been
additional functions, like attracting potential mates) was to
catch the rays of the sun. This would allow their possessors to
warm up quickly in the morning, and get on with the day's business
earlier than their cold-blooded cousins. They would have also
been able to stay more active during the day, and radiate away
heat more effectively on hot days. Some paleontologists believe
that these reptiles with solar panels may also have had systems
that shut down most circulation from the sails during cooler periods,
avoiding heat loss.
The sail system may have been just one
of the ways that reptiles employed in attempting to regulate their
body temperatures. In addition to the lineages that led to today's
reptiles -- snakes, lizards, alligators and crocodiles, and turtles
-- one highly successful reptile group -- the therapsids -- eventually
led to the mammals. The therapsids were so successful, in fact,
that they are estimated to have left behind about 90% of terrestrial
vertebrate fossils in the Late Permian. Huge herds of these animals
probably browsed the vegetation of southern Africa and elsewhere
in Gondwana.
One key to their success may have been
their thermoregulatory ability, the ability to regulate their
body temperatures. Whereas reptiles have scaly skins, which provide
little insulation, some therapsids may have evolved body coverings
of hair. Though we do not have evidence they wore fur coats, some
therapsid fossils do exhibit snout pits, dimples in facial bones
that may have accommodated those particularly sturdy hairs we
know as whiskers. If these therapsids had whiskers, they likely
had some body hair as well. Later therapsids may also have been
developing systems of internal thermoregulation, akin to mammalian
warm-bloodedness.
While the mammal-like reptiles were thriving,
other lines of reptiles were also evolving. Some of these reptiles
would eventually return to the seas, and dominate the Mesozoic
oceans as plesiosaurs and ichthyosaurs. Another reptile line would
lead to the large and fearsome carnivorous monitor lizards of
today's world, and also to the mosasaurs, another major group
of large Mesozoic marine reptiles. All of these marine reptiles
-- plesiosaurs, ichthyosaurs, and mosasaurs -- would meet their
fates at or near the end of the Cretaceous.
The last reptile lineage worth noting
here was that which eventually produced the crocodiles and alligators.
This reptile line would lead, in the Late Permian and Triassic,
first to the Archosauromorphs -- literally, the ruling lizard
forms -- and then to the Archosaurs, the "ruling lizards"
themselves. This descendants of this line of reptiles would take
over the position of dominant terrestrial vertebrates from the
mammal-like reptiles during the latter part of the Triassic. They
would also put at risk those mammal-like reptile lineages that
led, hundreds of millions of years later, to ourselves and all
modern mammals. These archosaur descendants were literally the
terrible lizards, the dinosaurs. They would dominate the land
for over a hundred and thirty million years.
In the two Permian oceans, Panthalassa
and the Tethys, fish and sharks were the dominant vertebrates.
Whales and dolphins -- marine mammals -- were hundreds of millions
of years in the future; after all, even true mammals were yet
to evolve. But the armored fish -- fish with great bony plates
on their heads -- of the earlier Paleozoic, particularly the Devonian,
were a distant memory. Though the fish and sharks of the Permian
oceans were not of types around today, their forms are familiar.
Not so the ammonites, which superficially
resemble today's chambered nautilus of the southwestern Pacific.
These often large, coiled mollusks were highly sophisticated organisms,
quite unlike their distant cousins, the clams and oysters. Like
their closer cousins, the octopi, ammonites were smart and stealthy
predators. They hunted and consumed fish, and competed with larger
fish for smaller prey.
On the shallow ocean floors of Permian
continental margins, trilobites were declining in number and variety
after thriving for hundreds of millions of years. Sponges were
plentiful, and there were carbonate reefs, though not the structurally
sophisticated reefs of the present. Two types of corals existed,
the cone-shaped rugose corals, and the flatter tabulate corals,
though neither were major reef constituents.
The single-celled marine organisms known
as foraminifers (referred to by most scientists as forams) were
flourishing, both in the water column and on the seafloor. These
organisms construct shells of calcium carbonate (like many sea
creatures, clams, for example) or build them by gluing together
grains of sand and bits of shell. Like amoebae, they have pseudopods
(literally, "false-feet": extensions of the cell body)
for locomotion and feeding, and consume both phytoplankton (plant-like
forms of plankton; they produce their own food) and zooplankton
(animal-like forms of plankton; they consume other organisms).
(Plankton is the name for organisms which passively drift in the
sea. Though they may have limited means of locomotion, for the
most part they are just carried along by ocean currents.) One
type of Permian foram, the fusulinids, was especially notable.
Perhaps with the cooperation of symbiotic algae, some fusulinids
reached the size of large grains of rice.
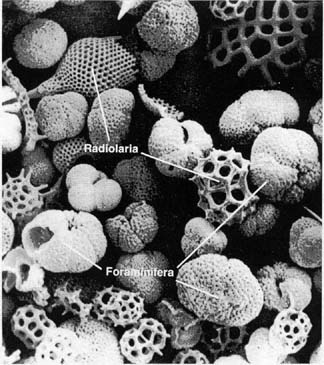 Present-day foraminifera. These tiny but complex single-celled organisms
have been around since the earliest Cambrian, about 540 million
years ago. Some forms float freely in the open ocean (that is,
they are pelagic); others live on the seafloor (that is, they
are benthic). Their complexity is revealed in the intricate skeletons
they build. Here the skeletons of the forams, as they are called,
are found together with the skeletons of other marine microorganisms,
the radiolaria. Radiolaria are also complex
single-celled organisms; they too have been around since Cambrian
times. Both forams and radiolaria do not manufacture their own
food, and thus are compelled to feed on other organisms or organic
debris. Radiolaria construct their
skeletons from silica (silicon dioxide, the same chemical composition
as glass); forams use various materials, including calcium carbonate.
The skeletons shown here are probably about
50 to 100 microns across. (A micron is a millionth of a meter,
or about 1/25,000ths of an inch.) (Thurman, 1993, Figure 4-5,
p. 89)
Present-day foraminifera. These tiny but complex single-celled organisms
have been around since the earliest Cambrian, about 540 million
years ago. Some forms float freely in the open ocean (that is,
they are pelagic); others live on the seafloor (that is, they
are benthic). Their complexity is revealed in the intricate skeletons
they build. Here the skeletons of the forams, as they are called,
are found together with the skeletons of other marine microorganisms,
the radiolaria. Radiolaria are also complex
single-celled organisms; they too have been around since Cambrian
times. Both forams and radiolaria do not manufacture their own
food, and thus are compelled to feed on other organisms or organic
debris. Radiolaria construct their
skeletons from silica (silicon dioxide, the same chemical composition
as glass); forams use various materials, including calcium carbonate.
The skeletons shown here are probably about
50 to 100 microns across. (A micron is a millionth of a meter,
or about 1/25,000ths of an inch.) (Thurman, 1993, Figure 4-5,
p. 89)
The oceanic food chain, like today's,
must have ultimately depended on photosynthetic organisms. Many
of these organisms were relatively simple and provided fodder
for the zooplankton. Lacking hard parts, they left no evidence
of their existence beyond possible organic residues on the ocean
floor. Other photosynthesizers were more complex, and constructed
minute shells of silica (glass). These photosynthesizers, the
diatoms, have been extremely important in Earth's oceans, and
continue so today.
The end-Permian Extinction
The magnitude of the extinction at the end of the Permian is unparalleled
in Earth's history. Although back in the dim reaches of time,
during the Precambrian, there may have been extinctions which
took out a higher percentage of individuals, and a higher percentage
of the then existing groups of organisms, the fossil record for
that time is extremely spare, and extinction events almost impossible
to recognize. During the eon of visible life (the Phanerozoic),
however, the geologic record has provided abundant fossils, making
it possible to identify major extinction events. Based on the
fossil evidence, therefore, the extinction at the end of the Permian
exceeds all other mass extinctions of the Phanerozoic.
(The end-Permian extinction may have
come in two stages, one several million years earlier than that
of the final catastrophe. Some scientists consequently believe
that the end-Permian was actually a kind of "double extinction,"
with a smaller but nonetheless major extinction coming perhaps
five million years before the ultimate event [see, for example,
Stanley and Yang, 1994; Racki, 2003]. This extinction came at
the end of the next-to-last geological interval [in this case
called an "age"] of the Permian, and is generally referred
to as the end-Guadalupian extinction. However, it is clear that
the devastation of this earlier event was not of the same scale
as that at the actual end of the Permian, and it is that extinction
which is our focus.)
According to some scientists (for example,
Stanley, 1987, p. 99), the forests and terrestrial plants of the
Permian are the group of organisms that best survived the end-Permian
extinction. Plants do tend to be more resistant to extinction
than other organisms, partly because they reproduce by means of
spores and seeds, which can survive long periods of unfavorable
climate conditions like drought. Plants also frequently are able
to reproduce from roots or subsurface stems and tubers, which
because of their location underground enjoy some protection from
events on the surface. Consequently, terrestrial plants have been
considered as relatively invulnerable to mass extinction (Knoll,
1984).
But terrestrial plants seem to have been
hard hit at the end of the Permian, according to some systematic
surveys of plant groups before and after the extinction. Over
an extended period of millions of years, the number of terrestrial
plant families dropped almost 50%, and the total number of species
fell about 20% (Knoll, 1984, Figures 1 and 2). This represents
the only significant loss of plant diversity in the entire Phanerozoic
(there was a lesser loss in the Late Devonian).
More detailed studies show that many
specific plants did become extinct at the end of the Permian;
these included, most notably, the broad-leafed, small- to medium-sized
tree Glossopteris, which had been ubiquitous throughout
Gondwana. Plants related to and associated with Glossopteris
(together called the Glossopteris flora) disappeared from
many parts of the world. In North China, a pteridophyte (fern,
horsetail and lycopod) flora replaced the conifers, while in South
China the number of species of spores and pollen fell by almost
one-half. This pattern was apparently worldwide, with similar
changes reported from Greenland, Poland, Hungary, Pakistan, Madagascar,
and Australia. The Early Triassic flora that replaced that of
the Permian included far fewer species (Yang, 1992).
 |
Glossopteris. This tree
and its close relatives were dominant members of Permian forests,
but they were devastated by the end-Permian extinction. Nonetheless,
some species did recover, and Glosssopteris trees were
present in the Triassic, and some may have survived into the
Jurassic. Some Glosssopteris trees stood about 4 meters
(13 feet) tall. Artists' renderings of
this tree vary considerably, however: Compare this drawing to
that in Appendix 2. Part of the difference in the depictions
can undoubtedly be traced to the fact that different parts of
the tree (leaves, pieces of branch, trunks) are found separately,
and the artist must reconstruct how they were actually assembled.
(Stewart and Rothwell, 1993, Figure 26.6, p. 372) |
In the Sydney area of southeastern Australia,
the plant extinction seems to have been particularly devastating:
97% of the Late Permian leaf species died off (Retallack, 1995).
Though spore data from this area indicates that a higher percentage
of plants may have survived than is indicated by the leaf fossils,
it is clear that the change in vegetation was sudden and profound.
Within only a short time, perhaps only a few thousand years, the
Glossopteris flora had been replaced by an Early Triassic
flora (Retallack, 1995). And there is no question that end-Permian
plant mortality must have been high: fungi, which live on dead
plant matter, did quite well at the time of the extinction.
We know this from the sudden increase
in the numbers of fungal spores preserved in the fossil record.
In fact, more than 95% of the spores found by one study at the
boundary between the Permian and Triassic Periods are fungal.
This "fungal spike" was worldwide in extent: similar
evidence has been found in North America, Greenland, Europe, Asia,
East Africa, Madacascar, and Australia (Eshet, 1995). The fungal
spores are found in every type of formerly aquatic environment
-- marine, lake, river -- an additional indication of how widespread
the fungal spike was (Visscher, 1996). Though plants are presumed
not as vulnerable to mass extinction events as other organisms,
they are also presumed to be more vulnerable to changes in climate.
Being rooted, they cannot escape when ecological conditions undergo
rapid and/or substantial shifts (Knoll, 1984).
Terrestrial animals, by contrast, suffered
extreme losses at the end of the Permian. Huge numbers of amphibians
and reptiles died off. (Mammals and birds, remember, had not yet
evolved.) More specifically, about three-quarters of the amphibian
and reptile families became extinct (Vickers-Rich and Rich, 1993,
p. 105). A family is a biological group containing many species,
which in turn may be composed of millions of individuals. (One
modern bird species contains only three individuals: this species
is one of many on their way to extinction in today's world. Other
species, such as those of bacteria, contain uncountable numbers.
Mammal species, on average, have about 40,000 breeding individuals,
though obviously this average includes some species with huge
numbers of individuals, such as humans or field mice, as well
as others which have far fewer, like tigers or polar bears.) For
a biological family to go extinct, every individual member of
every species in that family must die off.
Vertebrates like amphibians and reptiles
have hard parts like skeletons and teeth which make for good fossils.
Most other land animals have no such durable parts. Insects, for
example, have external skeletons built of proteins, which generally
do not survive for long lengths of time. The insect fossil record
is therefore quite incomplete, as is the fossil record for most
land invertebrates (creatures without backbones). Worms, slugs,
and other less familiar invertebrates leave even fewer traces
than insects. As a result, it is quite difficult to provide an
estimate of the losses suffered by these organisms at the end
of the Permian. There is no reason to suspect, however, that they
fared any better than other land animals.
We do have a better understanding of
the losses sustained by marine organisms, at least those which
have hard parts and fossilize well. (In addition, the constant
supply of sediment buries dead organisms and thereby helps preserve
them.) Fish and sharks apparently suffered only minor losses:
only about 10% of marine vertebrate families died off (Sepkoski,
1982). But among the marine invertebrates, the end-Permian devastation
was comparable to that of the land vertebrates. It is estimated
that between 90 and 96% of marine invertebrate species, and over
50% of marine invertebrate families, went extinct (Sepkoski, 1986).
These marine extinctions hit forams, gastropod (snail-like) and
bivalve mollusks, and echinoids (sea urchins, starfish, and their
relatives) hard.
Even harder hit were the rugose and tabulate
corals, which disappeared forever (though some believe that the
tabulate corals went earlier in the Permian; see Boardman, 1987),
ammonites, brachiopods (which superficially resemble clams but
are vastly different), and bryozoans (colonies of organisms with
lacy or platy skeletons). One arthropod group (the group that
includes crabs, lobsters, and all insects) that had been very
successful for more than two hundred million years, the eurypterids,
also met its demise. Another extremely successful group, the blastoids,
distant relatives of the starfish and the sea urchin, similarly
came to its end. Closer cousins of the blastoids, the crinoids,
many of which had stalks to lift them off the sea floor and allowed
them to filter-feed in higher currents, survived the extinction
with heavy losses (stalked crinoids are known as sea lilies, because
they superficially resemble lilies, but they are animals, not
plants). Only small groups of ammonites and bryozoans made it
across the boundary between the Permian and Triassic Periods.
In addition, the numbers of acritarchs -- enigmatic marine microfossils
with thick organic coats that are probably algal spores or cysts,
and are considered an "opportunistic survivor species"
-- increased dramatically (Retallack, 1995; Eshet, 1995).
Marine organisms that were attached to
or lived on the seafloor, including corals, bryozoans, echinoderms,
and certain brachiopods (articulate brachiopods), fared poorly
in the end-Permian extinction. These organisms, many of which
are also characterized by heavy carbonate skeletons, weak circulatory
systems and low metabolic rates, lost a high percentage of their
members (Knoll, 1996). Other marine organisms, with higher rates
of metabolism, stronger circulatory systems, and gills for better
absorption of dissolved oceanic gases, had far fewer losses. The
survival rates of these organisms -- mollusks, crustaceans, vertebrates
and exotic creatures like sea squirts -- in contrast to the low
survival rates of the others, has been attributed to their resistance
to poisoning by carbon dioxide, which is called hypercapnia (Knoll,
1996).
If this extinction pattern and its suggested
cause is correct, then end-Permian ocean waters did not merely
lose their oxygen, as has been suggested by numerous scientists,
but confronted unusually high carbon dioxide concentrations as
well. But carbon dioxide -- proposed, in this case, to have come
from ocean stagnation -- may not offer a unique solution to the
observed extinction pattern. Methane would also have disrupted
metabolic processes, and would have been rapidly converted by
chemical reactions in the oceans to carbon dioxide itself.
Although paleontologists can examine
the fossils of organisms that possessed hard parts, others without
such hard parts would have left behind no fossils, except possibly
biochemical traces in the sediments. However, these organisms
-- the cyanobacteria (formerly known as blue-green algae, though
they are not actually algae) -- are among the most important of
marine organisms both because they conduct photosynthesis, providing
the atmosphere with a good portion of its oxygen, and because
they are a major food source for innumerable other creatures.
While paleontologists are using increasingly
sophisticated techniques to identify such biochemical traces or
"molecular fossils" (also referred to as biomarkers),
we currently have no way of assessing what types of cyanobacteria
inhabited Permian oceans, or how they might have fared during
the end-Permian extinction event. This inability to obtain data
is most unfortunate, because cyanobacteria are likely to have
produced much of the Permian world's oxygen and used much of its
carbon dioxide, just as they do in today's oceans. Obviously,
a significant extinction among the cyanobacteria would have greatly
compromised primary productivity in the oceans, and been a major
factor in the extinction of those many other marine organisms
which depended on them, directly or indirectly, for food.
Other phytoplankton (some of which do leave
hard parts or other traces of their presence) were indeed apparently
badly hit in the end-Permian extinction (Payne, 2004).
If cyanobacteria and other phytoplankton
had been so compromised, three additional consequences would have
followed. Phytoplankton employ carbon dioxide in the process of
photosynthesis; with less photosynthesis going on, atmospheric
carbon dioxide levels would have risen even further. In fact,
it has been estimated that if all phytoplankton died off, atmospheric
carbon dioxide would have increased between 150 and 200 ppmv (parts
per million by volume)(Falkowski, 2000). This extra amount of
carbon dioxide would be negligible in a world where carbon dioxide
levels were already high, merely compounding an existing problem.
In addition, certain phytoplankton (for
example, Silicibacter pomeroyi) release into the atmosphere a
compound known as DMS (dimethyl sulfide: CH¸3SCH¸3.
According to Andrew Johnston of the University of East Anglia
in England, it is DMS which provides the distinctive smell of
the sea.) DMS is an extremely important cloud nucleating agent,
meaning that it helps clouds form by facilitating the condensation
of water vapor. (When DMS is oxidized, it releases its sulfur
as sulfur dioxide, SO¸2. The sulfur dioxide is converted
to sulfuric acid, H¸2SO¸4, which strongly attracts
water.) Fewer DMS-producing phytoplankton translate into reduced
DMS production; that in turn results in fewer clouds. In ocean
areas, decreased cloud cover would have meant greater absorption
of solar radiation by the dark waters, and therefore would have
been another source of global warming. (Generally, however, the
role of clouds themselves in the warming or cooling of the planet
is still in dispute, and appears to vary depending on factors
like the type of cloud, its altitude, droplet size, and so on.)
Finally, a major reduction of phytoplankton
would have reduced the influx of oxygen to the atmosphere. Oxygen
is a chemically active gas and readily combines with other elements
and compounds. Without free oxygen continually entering the atmosphere
after being produced by photosynthetic organisms, free oxygen
would decrease and eventually disappear from the atmosphere. If
phytoplankton -- or terrestrial green plants, or both -- suffered
significant extinction at the end of the Permian, therefore, the
supply of oxygen to the atmosphere would have been reduced, and
many aerobic organisms would have felt serious consequences.
|
Biological Classification
In assessing the damage caused by mass extinctions to the biological
world, paleontologists quantify the extinction's impact on various
groups of organisms. These groups are classified according to
how inclusive they are, just as for postal addresses. A postal
address includes the first name of the individual recipient,
the recipient's last name, an address number and the name of
a street, a city, state, and country. Moving up such a hierarchy
from the individual to the country involves ever more inclusive
groupings. Thus, there are many address numbers for most streets,
many streets in a city, many cities in a state.
Biological classification
involves the same sort of hierarchies. It is organized in the
following way:
| Domain |
|
|
|
|
|
|
|
| |
Kingdom |
|
|
|
|
|
|
| |
|
Phylum |
|
|
|
|
|
| |
|
|
Class |
|
|
|
|
| |
|
|
|
Order |
|
|
|
| |
|
|
|
|
Family |
|
|
| |
|
|
|
|
|
Genus |
|
| |
|
|
|
|
|
|
Species |
Each species belongs
to a genus, each genus to a family, and so on. Every species
has its own designation, consisting of two names, just like many
humans. Joe is an individual's name, for example, and Smith tells
us he is related to other Smiths. Similarly (although the order
of the individual name and the larger group are reversed), the
name Homo sapiens tells us that the species being referred to
is the species "sapiens," or wise, of the genus "Homo,"
or man. Though not part of its name, Homo is a primate, a mammal,
an animal, and a eukaryote (organisms with large, complex cells),
in this manner (Latin names are typically used):
| Eukaryota (domain) |
|
|
|
|
|
|
|
| |
Animalia (kingdom) |
|
|
|
|
|
|
| |
|
Chordata (phylum) |
|
|
|
|
|
| |
|
|
Mammalia (class) |
|
|
|
|
| |
|
|
|
Primates (order) |
|
|
|
| |
|
|
|
|
Homin- oidea (family) |
|
|
| |
|
|
|
|
|
Homo (genus) |
|
| |
|
|
|
|
|
|
sapiens (species) |
(Modified from Margulis and Schwartz, 1982, p. 3)
|
Each group in a biological
classification is made up of organisms that share particular attributes.
Thus, mammals are grouped together because they possess hair,
mammary glands, and other features. Human beings (Homo sapiens)
share these features with all other mammals, such as cats and
dogs, monkeys, mice, zebras and whales. Therefore humans are mammals
as well. Human beings do not possess scales or feathers, and therefore
are not reptiles (which have scales) or birds (which have feathers).
But, like reptiles and birds, we do have backbones and spinal
cords, which make us all -- human beings, other mammals, reptiles
and birds, together fish and amphibians and some other unfamiliar
organisms -- chordates. In addition, of course, we human beings
have our own special attributes which distinguish us from other
creatures.
When evaluating mass
extinctions, paleontologists often employ the higher groupings
(groupings are called taxa) to get the best sense of how various
organisms have been affected. Thus extinction compilations, though
they often indicate how many species went extinct, also frequently
include the impact at the family level. This helps provide a better
sense of just how badly a given group of organisms was hit.
APPENDIX
2: A GALLERY OF PERMIAN-TRIASSIC ORGANISMS
Summary of Biological Changes, Permian (and before) through
the Early Triassic
| |
Permian
(and before) |
end-Permian
extinction |
Early Triassic
and thereafter |
| Marine |
|
1/2 families
gone |
"Cosmopolitan"
biota (see below table for explanation.) |
| |
Fusulinids
(forams with
photosymbionts)
(found only in Tethys area by
Djulfian time) |
Gone |
|
| |
Other forams |
Hit hard |
|
| |
Reefs |
Decimated |
Recover, but
slowly and with different organisms |
| |
composed of
alcareous algae and calcareous sponges, bryozoans. |
Almost gone |
|
| |
Rugose corals
(found only in Tethys area by Djulfian time) |
Gone |
|
| |
Tabulate corals |
Gone |
|
| |
Scleractinian corals |
Survived |
Thrived after extinction |
| |
Bryozoans (lacy,
fanlike colonies
of organisms)
(found only in Tethys area by
Djulfian time) |
Hit hard |
Only one of the five Permian
orders survives but it thrives |
| |
Brachiopods |
Hit hard |
|
| |
Blastoids |
Gone |
|
| |
Crinoids |
Hit hard |
Recovered,
but never as plentiful as during Paleozoic |
| |
Bivalves
(mollusks) |
Hit, not badly |
|
| |
Gastropods |
Hit, not badly |
|
| |
Ammonites |
Hit hard |
|
| |
Trilobites
"quite uncommon" |
Trilobites
already gone? |
Major recovery.
Ammonites become significant predators in the Mesozoic seas |
| |
Eurypterids |
Gone |
|
| |
Bony fish and
sharks had replaced the placoderms (fish with bony armor plates) |
Hit, not badly |
Bony fish and
sharks thrive |
| Terrestrial
flora |
Lycopod (club
moss) trees replaced by drier climate trees, the gymnosperms: |
Hit |
Today's lycopods
are small but common plants |
| |
Cycads
Conifers (pines, firs, spruces) |
Survived extinction
fairly well |
Still around
Thrived in Mesozoic; lots around today |
| |
Ginkgoes |
Survived extinction fairly well |
Thrived in Mesozoic; still around |
| |
Glossopteris flora in Gondwana |
Gone |
Survived elsewhere,
but died out around the end of the Mesozoic |
| Terrestrial
fauna |
Big insects
in Carboniferous |
Gone; only
smaller insects left |
|
| |
During Late
Carboniferous,
Reptiles (amniote egg) evolve from: |
Hit hard
|
Crocodiles thrive.
Plenty of snakes, turtles, lizards still around
|
| |
Amphibians. |
Hit hard |
|
| |
More mammal-like
reptiles later in Permian: limbs under body, heterodonty
greater thermoregulation: higher body temps.
Pelycosaurs:
Edaphosaurus (herbivore) and
Dimetrodon, (carnivore) in Early Permian. Pelycosaurs in Laurasia
in Late Permian, but
therapsids (from Middle Permian) dominant on land
(Gondwana)
Richard Cowen (1990, p. 288) says 800 million fossil therapsids
estimated in Karroo Basin alone. |
Gone before end-Permian
|
Mammal and dinosaur ancestors survive
and thrive.
|
| |
Dicynodonts: highly diverse,
rat to cow size, such as: |
Some make it thru |
Survivors become quite
abundant: Lystrosaurus herds. |
| |
Dicynodon |
Gone |
|
Sources: Stanley, Extinctions,
1987.
(Stanley believes that extinction took about 10 million years,
and that cause was global cooling.)
Also Cowen, 1990; and Briggs and Crowther, 1990.
The Djulfian was the last stage of Permian (after the Guadalupian)
(In the Early Triassic, a large percentage
of the few survivor species had a very wide, that is, "cosmopolitan"
distribution. This is not surprising, because the survivors would
have had little competition, enabling them to occupy extensive
areas. Though cosmopolitan biotas are typically low in diversity,
many of the end-Permian survivors -- ammonites, therapsids, sharks
-- quickly diversified.)
In the mid-1990s, this is where our understanding
of the end-Permian extinction stood. All paleontologists recognized
that the riddle of mass extinctions generally had not been solved.
There might have been just a single cause of most mass extinctions,
or each may have had its own unique and unrepeatable cause.
At the University of California at Davis,
those of us who were graduate students in paleontology had our
own seminar, in which we discussed current articles in our field.
It was very much our own seminar, though nominally under faculty
guidance and advisorship. Faculty members were invited to attend,
but most had other, more pressing obligations. Occasionally, when
an article of particular relevance to a faculty member's own area
of specialization was to be discussed, that faculty member was
specifically invited to attend. But mostly we were on our own,
and suggested, selected and examined a wide variety of articles,
and had wide-ranging, sometimes quite animated, discussions.
Topics included the evolution of menstruation,
how fast a new species could evolve, why the tiny roundworm Caenorhabditis
elegans had been selected by biologists as a model organism for
investigating the process of biological development in animals,
oyster fossils from southern California mountains, why turtles
have their appendicular skeletons (that part of the skeleton which
includes pelvis, shoulders and limbs) inside their axial skeletons
(spine and ribs), the evolution of flight (a perennial favorite
of paleontologists), and so on.
At one of these noontime discussions,
at my suggestion, we looked at a newly published paper by Paul
Renne and his co-authors (1995) on the geologic dating of the
end Permian extinction and Siberian Traps volcanism. Renne is
a researcher at the Berkeley Geochronology Center, where they
attempt to decipher the mysteries of geologic time, by determining
the dates of origin for rocks and fossils.
Up to this point, the duration of the
end-Permian extinction had been unknown. But the Renne paper indicated
that the its duration was sufficiently short that it could be
considered an "event," rather than something more protracted.
The extreme shortness of the boundary event, however, severely
limited the possible causes. Impacts were rapid events, of course,
but the evidence then available suggested that there had been
no impact at the Permian-Triassic boundary. Additionally, more
exotic, and far more rare cosmic causes such as nearby supernova
explosions which could partly or completely sterilize the planet,
seemed to be ruled out because their nuclear residues were simply
not found by end-Permian investigators.
But if extraterrestrial causes could
be eliminated as possible contenders for the cause of the end-Permian
extinction, that left only terrestrial candidates. Moreover, any
real candidate would have had to do its work quickly, and leave
few traces. This was an unexpected, startling, and even frightening
possibility. Some terrestrial agent must have caused the greatest
extinction of the Phanerozoic. What possibly could be the killer
in our midst?
"A gas," I said. "Carbon
dioxide... No, methane."
I said methane for four reasons. First,
methane is an asphyxiating gas, and so can do similar kinds of
damage as carbon dioxide. Second, methane oxidizes to carbon dioxide
in just a short time (less than a decade), so with methane there
is a "two-fer": initially there is methane and later
carbon dioxide. With methane it is a case of, anything carbon
dioxide can do, methane can do better. Third, methane is a powerful
greenhouse gas, more powerful than carbon dioxide. Fourth, there's
a huge amount of methane sitting in the continental margins of
the world (presumably there was lots of it around in the Permian
as well), and, under the proper conditions, that methane could
have been released.
CONTINUE TO NEXT SECTION (Methane
and Methane Hydrates)
RETURN
TO CONTENTS